日本基礎生物學研究所植物環境應答研究部門的森田(寺尾)美代教授、西村嶽志助教和中村守貴特任研究員、福建農林大學的古谷將彥教授以及東京大學的平野良憲助教,與奈良先端科學技術大學院大學的箱嶋敏雄教授、基礎生物學研究所生命創成探究中心的加藤輝特任助教以及立命館大學的深尾陽一郎副教授等聯合研究,利用模式植物擬南芥發現了重力感受細胞中與重力方向的信號傳遞有關的新因子RLD。
植物的根部朝著地下、莖部朝著空中生長。這是植物感測重力(地心重力)方向發生的向地性(向地性)反應。此前雖然知道重力的方向通過根和莖的特殊細胞——重力感受細胞感知,植物向性通過控制生長素的傳輸實現對重力方向產生反應等,但一直不清楚重力感受細胞内的重力方向資訊傳遞機制的詳細情況。森田教授等人組成的研究團隊此前確認,名為LZY的蛋白質是向地性所需的物質。此次新發現了與LZY相輔作用的蛋白質RLD,同時確認,RLD也是與向地性有關的重要因子,RLD參與了生長素的傳輸調控。並提出生長素的傳輸調控分子機制,即LZY響應重力方向改變其在重力感受細胞内的位置,此時結合RLD並將其置於同一位置。相關研究成果已於2020年1月3日發布在國際學術期刊《Nature Communications》上。
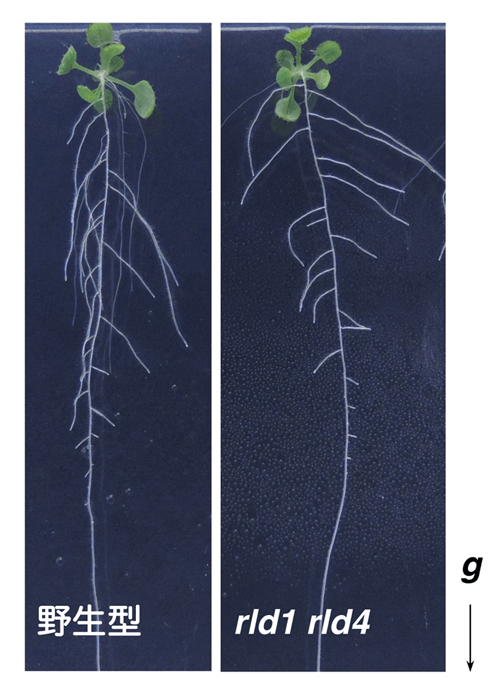
圖:通常,植物根部會感測重力的方向,表現出沿重力方向生長的向地性(左:野生型);但在rld1 rld4雙重突變體(右)中,向地性變弱,側根出現沿水平方向生長的傾向。
【研究背景】
向地性是植物通過感測重力方向配置身體各器官的重要環境響應特性之一,比如使根部朝著水分和營養成分豐富的地下生長,莖部朝著有利於進行光合作用和繁殖的上方生長。
在向性中,器官彎曲的機制是通過植激素生長素在器官内不均勻分布控制的。研究認為,根部向重力方向生長時,從地上方向通過中心部傳輸過來的生長素會均勻分布在根冠(圖1上左);當根部傾斜,重力方向改變時,生長素就會更多地分佈在重力側(圖1上右)。生長素濃度升高會抑制根部的細胞延伸,因此延長部域的根部會彎曲(圖1下)。這種器官内的生長素分配由生長素外輸載體PIN蛋白在細胞内的不均配置控制。
在根和莖等器官中都有感受重力方向變化(器官傾斜)的重力感受細胞。該細胞中含有的色素體會大量積聚澱粉形成澱粉體,其比重比細胞質高,會向重力方向沉降。已知這對識別重力方向至關重要。根部的重力感受細胞是位於根冠内的柱狀細胞(圖1下)。柱狀細胞的作用被認為是感受重力方向的變化並向重力側傳輸更多的生長素。不過,此前一直不清楚重力感受細胞内的重力信號傳遞機制。研究團隊此前利用模式植物擬南芥確認,植物特有的基因LZY(LAZY1-LIKE)家族在重力感受細胞内參與了重力刺激下生長素的分佈調控,但並不清楚其在重力感受細胞内具體負責什麼反應。
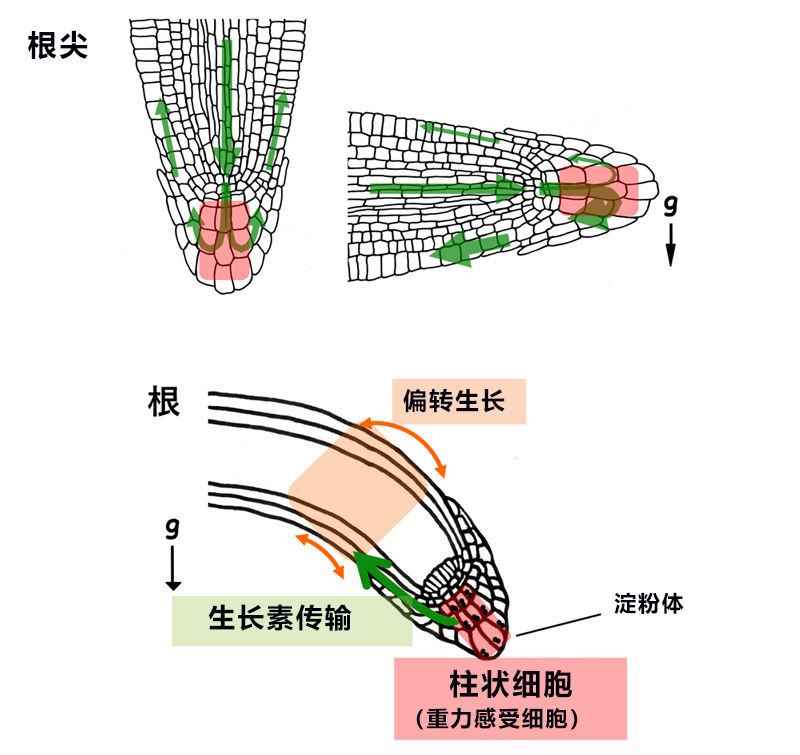
圖1:擬南芥根部的向地性反應
上:柱狀細胞(重力感受細胞)在根尖的位置(紅色)以及生長素的流動(綠色箭頭)。下:從重力感受細胞到器官偏轉生長的資訊動向。
【研究成果】
研究團隊首先確定了與LZY蛋白質相輔作用的因子,將具有類似結構的蛋白質命名為RLD(RCC1-like domain protein)1~4,並解析了其功能。解析發現:RLD1和RLD4在根部的重力感受細胞中發揮作用;rld1 rld4雙重突變體的向地性較弱,側根的生長接近水平方向;對重力進行響應的生長素傳輸出現異常(圖2)。由此可知,RLD與LZY在同一細胞中發揮同樣的作用。另外,利用rld1 rld2 rld3 rld4四重突變體進行解析確認,RLD可通過控制PIN蛋白來調節生長素傳輸。

圖2:擬南芥側根的生長方向
與野生型相比,rld1 rld4雙重突變體和lzy3突變體的側根傾向於水平生長。
研究確認,LZY與RLD是通過位於各自C末端的CCL域與BRX域相輔作用。這種結合起著重要作用,決定了這些蛋白質存在於細胞内的哪個位置。在培養細胞中,LZY存在於細胞膜,RLD存在於細胞質,二者分別存在於不同的位置。但當LZY和RLD在同一個細胞中表達時,RLD與LZY都存在於細胞膜中(圖3)。另外,二者共同存在於細胞膜依賴於CCL域與BRX域之間的相輔作用。此外,研究團隊純化得到了高純度的CCL-BRX蛋白質復合體,並在1000多種結晶條件下進行篩選後獲得了晶體(圖4左)。利用大型同步輻射設施(SPring-8,光量子工廠)對蛋白質晶體實施了X射線結構解析,成功確定了復合體的三維結構。BRX域的結構為首次報導。BRX域呈簡潔緊湊的形狀,能識別CCL獨具特色的β髮夾結構。另外,還在原子水平上明確了BRX與CCL通過形成分子間β片層而蠻力結合的機制(圖4右)。研究團隊根據該結構資訊,向LZY或RLD導入預計會抑制相輔作用的點突變。結果發現,兩種蛋白質之間的相輔作用消失,而且即使存在LZY,RLD也隻停留在細胞質中。另外,二者在重力感受細胞中的功能也失落。這表明,LZY將RLD引入細胞膜的性面質其在重力感受細胞中的功能很重要。
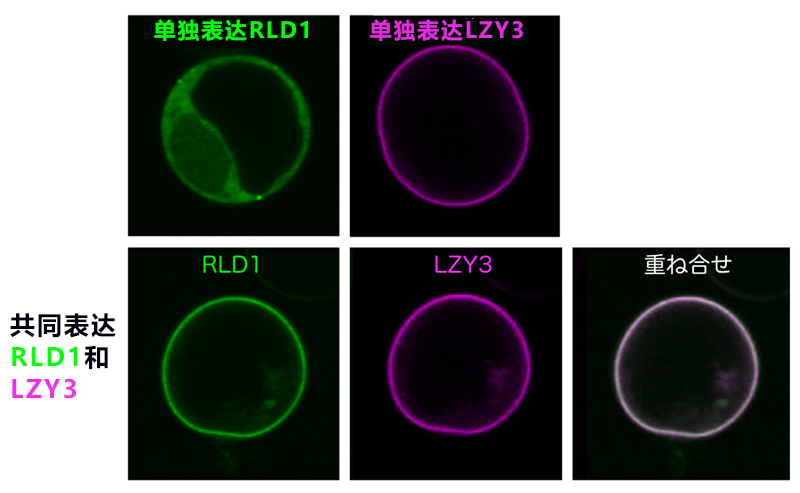
圖3:RLD通過LZY存在於細胞膜中
上:使培養細胞原生質體化,分別單獨表達RLD1和LZY3。RLD1存在於細胞質,LZY3存在於細胞膜中。下:同時表達RLD1和LZY3後,RLD1與LZY3一起存在於細胞膜中。
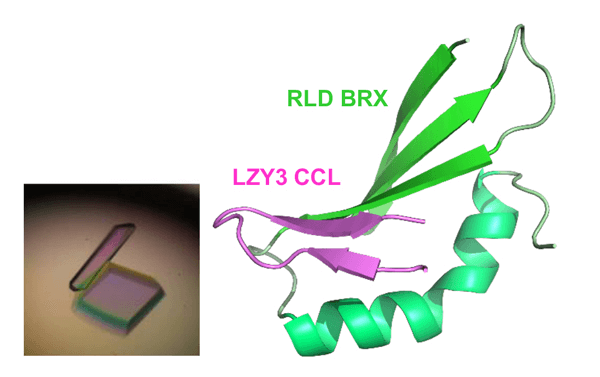
圖4. 蛋白復合體結構
左:LZY CCL域與RLD BRX域的蛋白復合體晶體
右:CCL-BRX復合體的X射線晶體結構
研究團隊分析了LZY和RLD在重力感受細胞内的工作機制。植物中的LZY蛋白質含量非常低,因此極難觀察。通過仔細挑選用於觀察的轉化子,並使固定的側根透明化,最終在作為重力感受細胞的柱狀細胞中成功擷取了微量的LZY蛋白質。有趣的是,LZY集中存在於重力側的細胞膜;旋轉180度並進行重力刺激30分鐘後,其出現在了新的重力側的細胞膜中(圖5)。此時,澱粉體也已經沉降到新的重力側。RLD通常在柱狀細胞内幾乎隻出現於細胞質中;同樣實施重力刺激60分鐘後,RLD也出現在了新的重力側的細胞膜中。在lzy突變體中則未觀察到這種現象。不過,LZY集中存在於重力側的現象在rld突變體中也被觀察到。另外,研究團隊調查在柱狀細胞中發揮作用的PIN3蛋白在根冠中的分佈情況時發現,在實施重力刺激5小時後野生型集中分佈於重力側,而rld1 rld4突變體中這種集中分佈大為減少。此外,在重力感受細胞中的人為過量表達LZY時,LZY會均勻分布於整個細胞膜中;在這種情況下,RLD也均勻分布在整個細胞膜中。此時,側根的生長方向會分散,由此可見,適度調節LZY蛋白質的量,並使其保持偏向於重力側的分佈對向地性至關重要。
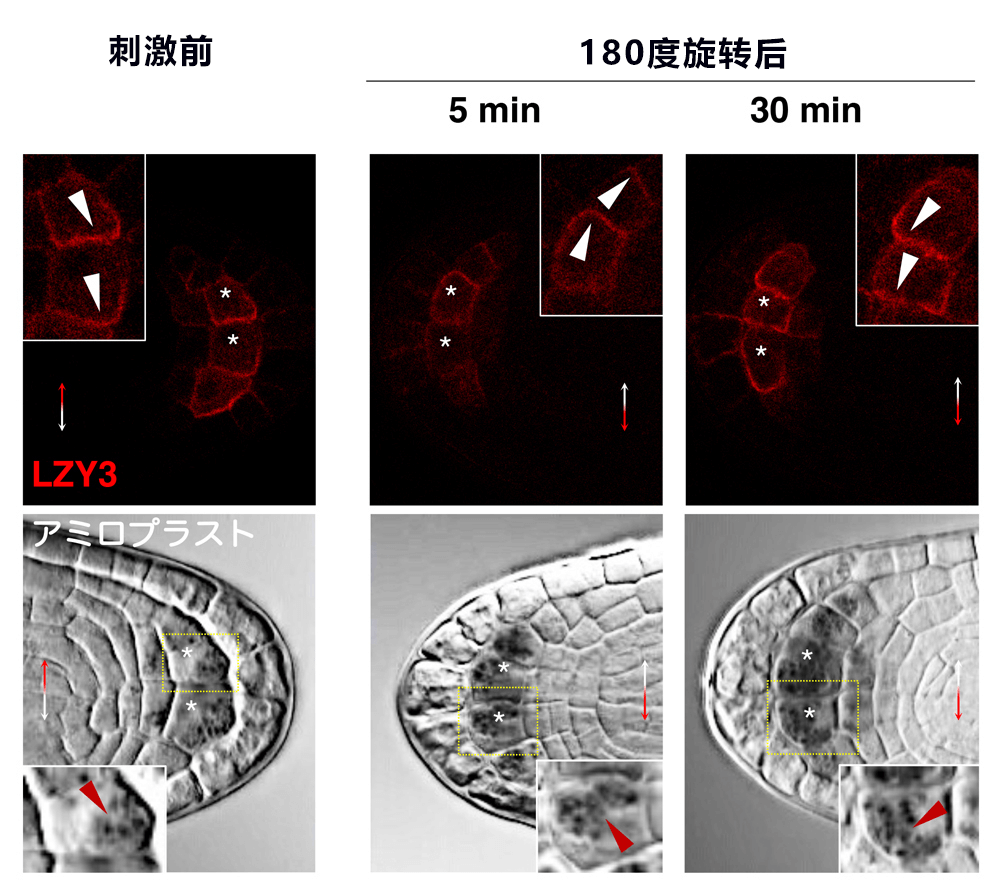
圖5:LZY3存在於重力方向側的細胞膜中
上:固定幼側根並使其透明化後觀察了LZY3。實施刺激前,LZY3集中分佈在重力側的細胞膜中。旋轉180度並實施重力刺激,5分鐘後,LZY3仍位於原來的重力方向,但30分鐘後則集中分佈在新的重力方向。下:澱粉體在5分鐘後幾乎沒移動,但30分鐘後移動到了新的重力方向。*:中央柱狀細胞,白色箭頭:LZY3的不均勻分布,紅色箭頭:澱粉體。
研究團隊根據以上結果提出了下述重力信號傳遞機制:在側根柱狀細胞中,①LZY基於澱粉體的位置資訊集中分佈在重力側的細胞膜中;②RLD根據CCL-BRX間的相輔作用從細胞質向重力側的細胞膜移動;③基於RLD的控制會使PIN蛋白向重力側集中;④更多地向重力側傳輸生長素(圖6)。
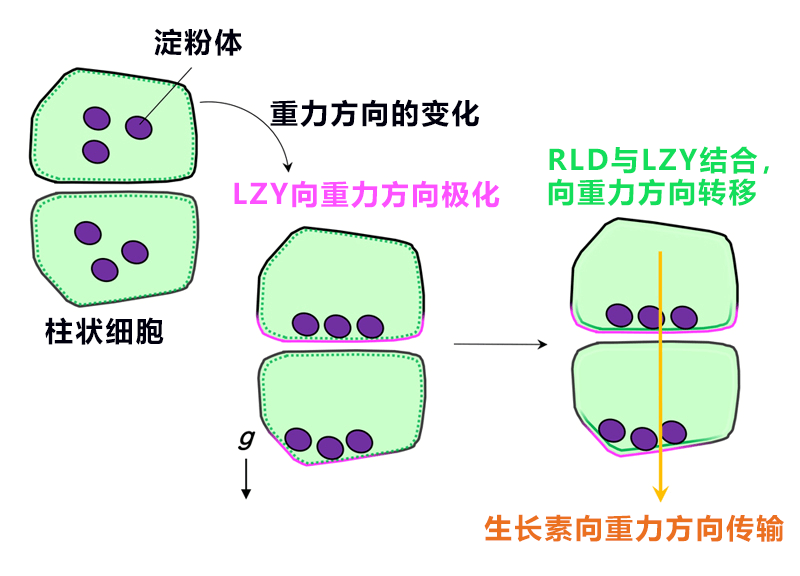
圖6:側根柱狀細胞的重力信號傳遞
【未來展望】
向地性還參與側枝和側根的生長角度控制,是通過植物器官的空間配置來控制株型(即枝條組態、根系組態)的主導因素之一,是育種的重要表徵。雖然在農業上屬於重要表徵,但始終不太瞭解其分子機制。此次,通過解析RLD的表達與LZY之間的功能性關係,具體明確了部分分子機制,比如澱粉體沉降後如何向重力側傳遞資訊。本次研究取得的成果將大大促進對向地性中的重力信號傳遞分子機制的理解。研究團隊的森田教授表示:「今後打算進一步調查LZY集中存在於重力側細胞膜中的機制,以查清植物的重力方向識別機制。另外,通過進一步詳細解析RLD對PIN的調節,希望能瞭解對植物至關重要的生長素的傳輸調控機制」。
論文資訊
題目:Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control
期刊:《Nature Communications》
DOI:https://doi.org/10.1038/s41467-019-13729-7
文:JST客觀日本編輯部翻譯編譯