奈良先端科學技術大學院大學先端科學技術研究科白川一助教、山口暢俊副教授、伊藤壽朗教授,以及京都大學研究生院生命科學研究科的山岡尚平副教授和河內孝之教授等人的研究團隊宣佈,氣孔形成因子在十字花科植物中被「轉用」於負責免疫防禦細胞的形成中。針對開啟和關閉植物葉片氣孔的保衛細胞和負責免疫防禦的白芥子酶細胞這2種不同功能的細胞分化,本研究成功鑑定出一種參與相應細胞基因表現的新型轉錄因子「WASABI MAKER(WSB)」。這一發現有望應用於包括改變作物中辛辣成分數量和質量的技術,以及開發能高效吸收二氧化碳的氣孔優化植物。相關研究成果已發表在國際學術期刊《Nature Plants》的2月24日刊上。
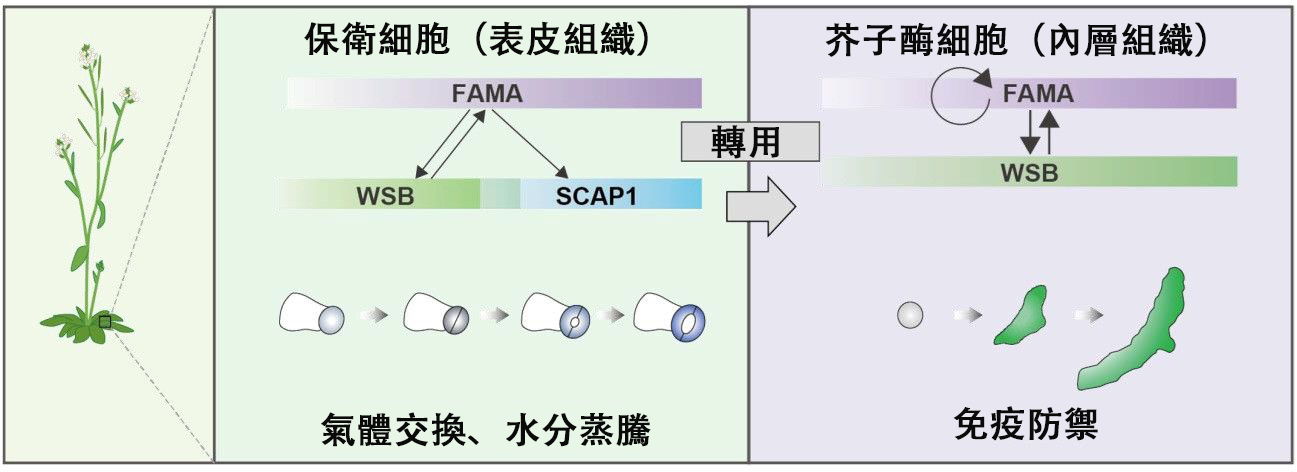
圖1研究成果示意圖(供圖:奈良先端科學技術大學院大學白川一助教)
植物能夠產生動物所沒有的特殊細胞,但它們的基因,以及在有限的基因數量中產生這些特殊細胞的機制尚未明確。
此次研究團隊關注了2種特化的細胞,即「保衛細胞」和「白芥子酶細胞」。保衛細胞分佈在幾乎所有陸生植物葉片的表面,負責氣體交換和水分蒸騰。白芥子酶細胞是十字花科植物才會產生的特殊細胞,當白芥子酶細胞中含有的一種名為白芥子酶的酶與受質硫代葡萄糖苷(芥子油苷)發生反應時間,就會產生揮發性的驅避物質異硫氰酸酯,從而起到免疫防禦作用。硫代葡萄糖苷積聚在不同於白芥子酶細胞的另一種特化細胞(S細胞)中,只有當細胞受到外部侵害遭到破壞時,才會產生高反應性的驅避物質。異硫氰酸酯一方面作為免疫防禦物質對抗植食性昆蟲和細菌,另一方面也是山葵和辣根辛辣成分的來源。
此前的研究已經表明,轉錄因子「FAMA」是從生物設計圖DNA中產生其副本mRNA的作用因子,是保衛細胞和白芥子酶細胞分化所必需的。另一方面,它的原理並不明確,儘管它是兩種細胞分化所必需的,但人們並不清楚它是如何使這兩種細胞分化出不同功能的。
對此,通過全面基因表現分析,研究團隊鑑定出了受下游控制因子FAMA顯著影響的候選基因體。在FAMA誘導後立即受到影響的約600個基因中,通過與之前報告的全面基因表現分析數據相結合,最終鎖定並鑑定出了1個基因,即WASABI MAKER轉錄因子(WSB)。
通過染色質免疫沉澱法和報告基因檢測,研究團隊確認了FAMA直接與啟動WSB表達的位點(啟動子)結合,並對其表達進行控制。
此外,研究團隊還揭示了WSB對FAMA的積極反饋,以及FAMA和WSB在功能上的相互依賴。
研究團隊進一步通過基因編輯技術生成了WSB功能缺失突變體,並對其進行了白芥子酶細胞分化檢測,但與FAMA突變體一樣,WSB突變體中也未能觀察到白芥子酶細胞的形成。
由此可以推斷,FAMA是通過WSB來促進白芥子酶細胞分化的。
此外,WSB突變體的保衛細胞並不像FAMA突變體的保衛細胞那樣異常,這促使研究團隊開始尋找FAMA的下游靶標因子。研究發現,SCAP1轉錄因子是保衛細胞獨特性FAMA的直接靶標因子,與WSB突變體類似,SCAP1突變體也形成了正常的保衛細胞。與此相反,在WSB和SCAP1這2個轉錄因子被破壞的突變體中,正常保衛細胞的形成比例僅佔野生型的3%左右。
由此可知,FAMA是通過WSB和SCAP1來促進保衛細胞分化的。
通過譜系學樹分析,WSB基因在進化過程中被保守了多少,發現廣泛保守在被子植物中。研究團隊認為,WSB在被子植物的保衛細胞分化過程中發揮著廣泛的作用,在十字花科植物出現時,還獲得了製造白芥子酶細胞的功能。
在十字花科植物的WSB基因為了形成白芥子酶細胞而獲得了新功能這一假設的基礎上進行實驗發現,WSB通過抑制SCAP1,阻止了白芥子酶細胞獲得保衛細胞樣特徵。研究還發現,WSB對SCAP1的抑制調控僅在白芥子酶細胞分化過程中發揮作用。
研究表明,FAMA-WSB-SCAP1系統在保衛細胞分化中起作用,而其轉錄網路的簡化版本FAMA-WSB系統則被轉用至白芥子酶細胞分化中。
白川助教表示,「我們在研究十字花科植物的免疫防衛機制時,關注到了一種非常特殊的細胞,並意外地發現了它們與‘氣孔形成機制’之間的深度聯繫。氣孔形成是植物科學領域最熱門的話題之一,因此這也是一項極具影響力的發現。未來,我們希望能闡明進化過程中氣孔形成因子的轉用及獲得新功能的分子機制」。
原文:《科學新聞》
翻譯:JST客觀日本編輯部
【論文資訊】
期刊:Nature Plants
論文:Co-option and neofunctionalization of stomatal executors for defense against herbivores in Brassicales
DOI:10.1038/s41477-025-01921-1